

Видео:Площадь сферы внутри цилиндра. Поверхностный интегралСкачать

Центральный цилиндр). Анатомия корня. Первичное строение корня (эпиблема, первичная кора,
Анатомия корня. Первичное строение корня (эпиблема, первичная кора,
Первичное строение корня — дифференциация тканей корня происходит в зоне всасывания. По происхождению это первичные ткани, так как они образуются из первичной меристемы зоны роста. Поэтому микроскопическое строение корня в зоне всасывания называют первичным. При первичном строении в корне различают центральный цилиндр и первичную кору, покрытую одним слоем клеток с корневыми волосками — эпиблемой и ризодермой.
Клетки ризодермы вытянуты по длине корня. При их делении образуются два вида клеток: трихобласты, развивающие корневые волоски, и атрихобласты, выполняющие функции покровных клеток. В отличие от клеток эпидермы они тонкостенные и кутикулы не имеют. Расположены трихобласты поодиночке или группами. Корни, развивающиеся в воде, обычно не имеют корневых волосков, если же эти корни затем проникают в почву, волоски образуются в большом количестве. При отсутствии волосков вода проникает в корень через тонкие наружные стенки клеток.
Корневые волоски появляются в виде небольших выростов трихобластов. Рост волоска происходит у его верхушки. Благодаря образованию волосков общая поверхность всасывающей зоны увеличивается в 10 раз и более. Их длина 1. 2 мм. Корневые волоски недолговечны. Продолжительность их жизни не превышает 10. 20 дней. После их отмирания ризодерма постепенно сбрасывается. К этому времени подстилающий ее слой клеток первичной коры дифференцируется в защитный слой — экзодерму. Ее клетки плотно сомкнуты, после опадения ризодермы их стенки опробковевают. Нередко опробковевают и примыкающие к ней клетки первичной коры. Экзодерма функционально сходна с пробкой, но отличается от нее расположением клеток: таблитчатые клетки пробки, образуемые при тангентальных делениях клеток пробкового камбия (феллогена), распола-гаются на поперечных срезах правильными рядами, а клетки многослойной экзодермы, имеющие многоугольные очертания, — в шахматном порядке. В мощно развитой экзодерме нередко встречаются пропускные клетки с неопробковевшими стенками.
Остальная часть первичной коры — мезодерма, за исключением самого внутреннего слоя, дифференцирующегося в эндодерму, состоит из паренхимных клеток, наиболее плотно расположенных в наружных слоях. В средней и внутренней частях коры клетки мезодермы имеют более или менее округлые очертания, нередко самые внутренние клетки составляют радиальные ряды. Между клетками возникают межклетники, а у некоторых водных и болотных растений—довольно крупные воздухоносные полости.
Клетки коры снабжают ризодерму пластическими веществами и сами участвуют в поглощении и проведении веществ. Самый внутренний слой коры — эндодерма, которая выполняет роль барьера, контролирующего перемещение веществ из коры в центральный цилиндр и обратно.
Эндодерма состоит из плотно сомкнутых клеток, слегка вытянутых и почти квадратных в поперечном сечении. В молодых корнях ее клетки имеют пояски Каспари — участки стенок, характеризующиеся наличием веществ, химически сходных с суберином и лигнином. Пояски Каспари опоясывают поперечные и продольные радиальные стенки клеток посередине. Вещества, откладывающиеся в поясках Каспари, закрывают отверстия находящихся в этих местах плазмодесменных канальцев. У многих двудольных и голосеменных растений дифференциация эндодермы обычно заканчивается образованием поясков Каспари.
У однодольных растений, не имеющих вторичного утолщения, со временем эндодерма изменяется. Процесс опробковения распространяется на поверхность всех стенок, перед этим сильно утолщаются радиальные и внутренние тангентальные стенки, а наружные почти не утолщаются. В этих случаях говорят о подковообразных утолщениях. Утолщенные стенки клеток впоследствии одревесневают, протопласты отмирают. Некоторые клетки остаются живыми, тонкостенными, только с поясками Каспари, их называют пропускными. Они обеспечивают физиологическую связь между первичной корой и центральным цилиндром. Обычно пропускные клетки расположены против тяжей ксилемы.
Центральный цилиндр корня состоит из двух зон: перициклической и проводящей. В корнях некоторых растений внутреннюю часть центрального цилиндра составляет механическая ткань, или паренхима, но эта «сердцевина» не гомологична сердцевине стебля, так как слагающие ее ткани имеют прокамбиальное происхождение.
Перицикл может быть однородным и неоднородным. Он может быть однослойным и многослойным. Перицикл представляет собой меристему, так как он играет роль корнеродного слоя — в нем закладываются боковые корни, а у корнеотпрысковых растений — придаточные почки. У двудольных и голосеменных растений он участвует во вторичном утолщении корня, образуя феллоген и частично камбий. Клетки его долго сохраняют способность к делению. Выше зоны ветвления стенки клеток перицикла у некоторых растений одревесневают.
Первичные проводящие ткани корня составляют сложный проводящий пучок, в котором радиальные тяжи ксилемы чередуются с группами элементов флоэмы. Его образованию предшествует заложение прокамбия в виде центрального тяжа. Дифференциация клеток прокамбия в элементы протофлоэмы, а затем и протоксилемы начинается на периферии, в дальнейшем эти ткани развиваются центростремительно.
Если закладывается один тяж ксилемы и соответственно один тяж флоэмы, пучок называют монархным (такие пучки встречаются у некоторых папоротников), если по два тяжа — диархным, как у многих двудольных, у которых могут быть также три-, тетра- и пен- тархные пучки. Корням однодольных свойственны полиархные пучки.
В каждом радиальном тяже ксилемы внутрь от элементов протоксилемы дифференцируются более широкопросветные элементы метаксилемы.
Сформированный тяж ксилемы может быть довольно коротким (ирис), внутренняя часть прокамбия в этом случае дифференцируется в механическую ткань. У других растений (лука, тыквы) ксилема на поперечных срезах корней имеет звездчатые очертания, в самом центре корня находится наиболее широкопросветный сосуд метаксилемы, от него лучами отходят тяжи ксилемы, состоящие из элементов, диаметры которых постепенно уменьшаются от центра к периферии. У многих растений с полиархными пучками (злаки, осоки, пальмы) отдельные элементы метаксилемы могут быть разбросаны по всему поперечному сечению центрального цилиндра между паренхимными клетками или элементами механической ткани.

Первичная флоэма, как правило, состоит из тонкостенных элементов, лишь у некоторых растений (фасоль) развиваются протофлоэмные волокна.
Видео:11 класс. Геометрия. Объем цилиндра. 14.04.2020Скачать

Устройство современного двигателя
Видео:Объём цилиндраСкачать

Цилиндры двигателя
Цили́ндр двигателя внутреннего сгорания является рабочей камерой объемного вытеснения. Во время работы двигателя внутренние и наружные части цилиндров испытывают различный нагрев.
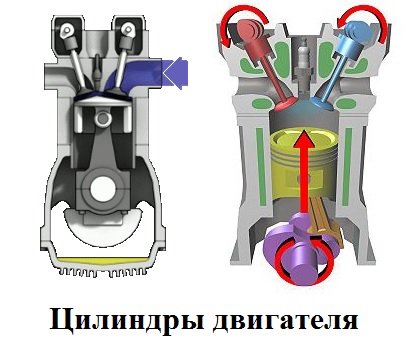
Внутренняя часть цилиндра — втулка или гильза цилиндра.
Наружная часть — рубашка двигателя.
Внутренняя поверхность втулки или гильзы цилиндра называется зеркалом. Зеркало это рабочая часть цилиндра, поэтому она подвергается специальной обработке (хонингование, хромирование, азотирование) и поэтому выбирают следующие типы материалов для гильз цилиндров. На зеркале цилиндра наносится специальный рельеф, который способствует снижению трения между поршнем, поршневыми кольцами и цилиндром, благодаря удерживанию моторного масла на стенках.
В современных двигателях внутреннюю поверхность цилиндров подвергают отбеливающему переплаву лазером, что способствует образованию белого чугуна высокой твердости. Ресурс таких цилиндров намного выше и не требует ремонтных размеров.
Гильзы цилиндров отливают из чугуна высокой прочности или специальных сталей. Иногда на алюминиевые гильзы цилиндров наносят гальваническое покрытие хромом.
В одноцилиндровом четырехтактном двигателе коленчатый вал вращается неравномерно, поэтому маховик должен обладать большим моментом инерции. В многоцилиндровом двигателе вращение коленчатого вала происходит равномернее, так как рабочие ходы в различных цилиндрах не совпадают друг с другом. Чем больше цилиндров имеет двигатель, тем равномернее вращается коленчатый вал. Нагрузка на детали кривошипно-шатунного механизма в многоцилиндровом двигателе изменяется более плавно, чем в одноцилиндровом.
Цилиндры двигателя могут быть расположены следующим образом: вертикально в один ряд – однорядные, двигателя автомобилей ВАЗ-2107 «Жигули», ГАЗ-52-04, ГАЗ-3102 «Волга» и др., под углом a к вертикали, двигатель автомобиля Москвич 2140; в два ряда V-образные, двигателя автомобилей ГАЗ-53А,ЗИЛ-130, КаМаз 5320 и др.
Видео:Внутреннее строение стебля. 7 класс.Скачать

Дефекты гильз цилиндров
Гильзы цилиндров изнашиваются вследствие трения между поршнем и зеркалом (внутренней стенкой цилиндра). Как правило повышенный износ может происходить вследствие таких причин:
— не достаточно масла на стенках цилиндров
-двигатель долго не работал, и все масло стекло в картер
-применение масла не соответствующей вязкости
— коррозия, возникает вследствии применения воды, как охлаждающей жидкости
-сколы, царапины возникают вследствие не правильного монтажа, демонтажа ( все действия по съемке гильз цилиндров нужно проводить согласно правил специальным съемником)
-при не правильной эксплуатации двигателя
Видео:Секретные похороны российских военных ПитербургСкачать

Методы обработки для устранения дефектов
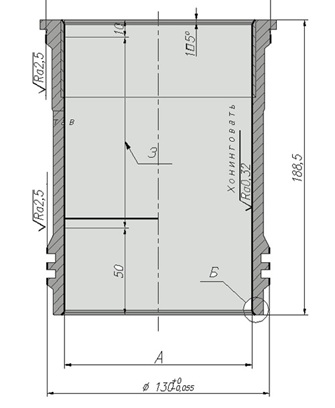
Дефекты устраняются такими методами обработки как: шлифовка, фрезировка, напыление, наплавка, хонингование.
Видео:Анатомия растений. Корень (Вислобоков Н. А.)Скачать

Хонингование
Хонингование — вид абразивной обработки материалов с применением хонинговальных головок (хонгов). В основном применяется для обработки внутренних цилиндрических отверстий (от 2 мм) путём совмещения вращательного и поступательно-возвратного движения хона с закреплёнными на нём раздвижными абразивными брусками с обильным орошением обрабатываемой поверхности смазочно-охлаждающей жидкостью. Один из видов чистовых и отделочных обработок резанием. Позволяет получить отверстие с отклонением от цилиндричности до 5 мкм и шероховатостью поверхности Ra=0.63÷0.04.
Обработка отверстий в различных деталях в том числе в деталях двигателя (отверстий блоков цилиндров, гильз цилиндров, отверстий кривошипной и поршневой головок шатунов, отверстий шестерен) и т. д. При обработке хонингованием обеспечивается стабильное получение точных отверстий и требуемых параметров шероховатости обработанной поверхности. Зеркало цилиндров должно иметь не совсем гладкую поверхность, так как масло будет стекать и не оставатся между парой трения, что будет приводить к износу, поэтому делается как бы меленькая насечка. В ней остаються частички масла , которые обеспечивают хорошую работу цилиндр-поршень и приводит к увеличению ресурса деталей.
Видео:Почему никто не знает об этой функции штангенциркуля?!Скачать

Центральный цилиндр).
Первичное строение корня — дифференциация тканей корня происходит в зоне всасывания. По происхождению это первичные ткани, так как они образуются из первичной меристемы зоны роста. Поэтому микроскопическое строение корня в зоне всасывания называют первичным. При первичном строении в корне различают центральный цилиндр и первичную кору, покрытую одним слоем клеток с корневыми волосками — эпиблемой и ризодермой.
Клетки ризодермы вытянуты по длине корня. При их делении образуются два вида клеток: трихобласты, развивающие корневые волоски, и атрихобласты, выполняющие функции покровных клеток. В отличие от клеток эпидермы они тонкостенные и кутикулы не имеют. Расположены трихобласты поодиночке или группами. Корни, развивающиеся в воде, обычно не имеют корневых волосков, если же эти корни затем проникают в почву, волоски образуются в большом количестве. При отсутствии волосков вода проникает в корень через тонкие наружные стенки клеток.
Корневые волоски появляются в виде небольших выростов трихобластов. Рост волоска происходит у его верхушки. Благодаря образованию волосков общая поверхность всасывающей зоны увеличивается в 10 раз и более. Их длина 1. 2 мм. Корневые волоски недолговечны. Продолжительность их жизни не превышает 10. 20 дней. После их отмирания ризодерма постепенно сбрасывается. К этому времени подстилающий ее слой клеток первичной коры дифференцируется в защитный слой — экзодерму. Ее клетки плотно сомкнуты, после опадения ризодермы их стенки опробковевают. Нередко опробковевают и примыкающие к ней клетки первичной коры. Экзодерма функционально сходна с пробкой, но отличается от нее расположением клеток: таблитчатые клетки пробки, образуемые при тангентальных делениях клеток пробкового камбия (феллогена), распола-гаются на поперечных срезах правильными рядами, а клетки многослойной экзодермы, имеющие многоугольные очертания, — в шахматном порядке. В мощно развитой экзодерме нередко встречаются пропускные клетки с неопробковевшими стенками.
Остальная часть первичной коры — мезодерма, за исключением самого внутреннего слоя, дифференцирующегося в эндодерму, состоит из паренхимных клеток, наиболее плотно расположенных в наружных слоях. В средней и внутренней частях коры клетки мезодермы имеют более или менее округлые очертания, нередко самые внутренние клетки составляют радиальные ряды. Между клетками возникают межклетники, а у некоторых водных и болотных растений—довольно крупные воздухоносные полости.
Читайте также: Прямой цилиндр это как
Клетки коры снабжают ризодерму пластическими веществами и сами участвуют в поглощении и проведении веществ. Самый внутренний слой коры — эндодерма, которая выполняет роль барьера, контролирующего перемещение веществ из коры в центральный цилиндр и обратно.
Эндодерма состоит из плотно сомкнутых клеток, слегка вытянутых и почти квадратных в поперечном сечении. В молодых корнях ее клетки имеют пояски Каспари — участки стенок, характеризующиеся наличием веществ, химически сходных с суберином и лигнином. Пояски Каспари опоясывают поперечные и продольные радиальные стенки клеток посередине. Вещества, откладывающиеся в поясках Каспари, закрывают отверстия находящихся в этих местах плазмодесменных канальцев. У многих двудольных и голосеменных растений дифференциация эндодермы обычно заканчивается образованием поясков Каспари.
У однодольных растений, не имеющих вторичного утолщения, со временем эндодерма изменяется. Процесс опробковения распространяется на поверхность всех стенок, перед этим сильно утолщаются радиальные и внутренние тангентальные стенки, а наружные почти не утолщаются. В этих случаях говорят о подковообразных утолщениях. Утолщенные стенки клеток впоследствии одревесневают, протопласты отмирают. Некоторые клетки остаются живыми, тонкостенными, только с поясками Каспари, их называют пропускными. Они обеспечивают физиологическую связь между первичной корой и центральным цилиндром. Обычно пропускные клетки расположены против тяжей ксилемы.
Центральный цилиндр корня состоит из двух зон: перициклической и проводящей. В корнях некоторых растений внутреннюю часть центрального цилиндра составляет механическая ткань, или паренхима, но эта «сердцевина» не гомологична сердцевине стебля, так как слагающие ее ткани имеют прокамбиальное происхождение.
Перицикл может быть однородным и неоднородным. Он может быть однослойным и многослойным. Перицикл представляет собой меристему, так как он играет роль корнеродного слоя — в нем закладываются боковые корни, а у корнеотпрысковых растений — придаточные почки. У двудольных и голосеменных растений он участвует во вторичном утолщении корня, образуя феллоген и частично камбий. Клетки его долго сохраняют способность к делению. Выше зоны ветвления стенки клеток перицикла у некоторых растений одревесневают.
Первичные проводящие ткани корня составляют сложный проводящий пучок, в котором радиальные тяжи ксилемы чередуются с группами элементов флоэмы. Его образованию предшествует заложение прокамбия в виде центрального тяжа. Дифференциация клеток прокамбия в элементы протофлоэмы, а затем и протоксилемы начинается на периферии, в дальнейшем эти ткани развиваются центростремительно.

Если закладывается один тяж ксилемы и соответственно один тяж флоэмы, пучок называют монархным (такие пучки встречаются у некоторых папоротников), если по два тяжа — диархным, как у многих двудольных, у которых могут быть также три-, тетра- и пен- тархные пучки. Корням однодольных свойственны полиархные пучки.
В каждом радиальном тяже ксилемы внутрь от элементов протоксилемы дифференцируются более широкопросветные элементы метаксилемы.
Сформированный тяж ксилемы может быть довольно коротким (ирис), внутренняя часть прокамбия в этом случае дифференцируется в механическую ткань. У других растений (лука, тыквы) ксилема на поперечных срезах корней имеет звездчатые очертания, в самом центре корня находится наиболее широкопросветный сосуд метаксилемы, от него лучами отходят тяжи ксилемы, состоящие из элементов, диаметры которых постепенно уменьшаются от центра к периферии. У многих растений с полиархными пучками (злаки, осоки, пальмы) отдельные элементы метаксилемы могут быть разбросаны по всему поперечному сечению центрального цилиндра между паренхимными клетками или элементами механической ткани.
Первичная флоэма, как правило, состоит из тонкостенных элементов, лишь у некоторых растений (фасоль) развиваются протофлоэмные волокна.
33.Типичный корень представляет собой подземный орган, присущий всем высшим растениям (кроме мхов). Корень служит для закрепления растения в почве, поглощения из почвы воды с растворенными в ней солями, в корне часто откладываются запасные продукты, корень участвует в синтезе органических веществ, служит для вегетативного размножения. Корень никогда не несет на себе листьев, поэтому по сравнению с внутренней структурой стебля у корня она относительно проста.
Корень по длине можно разделить на несколько участков, имеющих различное строение и выполняющих различные функции. Эти участки называют зонами корня. Выделяют корневой чехлик и следующие зоны: деления, растяжения, всасывания и проведения.
Дифференциация тканей корня происходит в зоне всасывания. По происхождению это первичные ткани, так как они образуются из первичной меристемы конуса нарастания. Поэтому микроскопическое строение корня в зоне всасывания называют первичным. У однодольных растений первичное строение сохраняется и в зоне проведения. Здесь лишь отсутствует самый поверхностный слой с корневыми волосками — ризодерма (эпиблема). Защитную функцию выполняет ниже лежащая ткань — экзодерма.
В первичном строении корня выделяют три части: ризодерму, первичную кору и осевой (центральный) цилиндр.
Строение ризодермы рассматривалось в теме «Покровные ткани».
На первичную кору приходится основная масса первичных тканей корня. Ее клетки накапливают крахмал и другие вещества. Эта ткань содержит многочисленные межклетники, имеющие значение для аэрации клеток корня. Наружные клетки первичной коры, лежащие непосредственно под ризодермой, называются экзодермой. Основная масса коры (мезодерма) образована паренхимными клетками. Самый внутренний слой носит название эндодермы. Это ряд плотно сомкнутых клеток (без межклетников).
Центральный или осевой цилиндр (стела) состоит из проводящих тканей, окруженных одним или несколькими слоями клеток — перициклом.
Внутренняя часть центрального цилиндра у большинства растений занимает сплошной тяж первичной ксилемы, дающий к перициклу выступы в виде ребер. Между ними размещаются тяжи первичной флоэмы.
У двудольных и голосеменных растений уже в раннем возрасте в центральном цилиндре корня между ксилемой и флоэмой появляется камбий, деятельность которого приводит к вторичным изменениям и в конечном итоге формируется вторичная структура корня. К центру камбий откладывает клетки вторичной ксилемы, а к периферии — клетки вторичной флоэмы. В результате деятельности камбия первичная флоэма оттесняется кнаружи, а первичная ксилема остается в центре корня.
Вслед за изменениями в центральном цилиндре корня происходят изменения в коровой части. Клетки перицикла начинают делиться по всей окружности, в результате чего возникает слой клеток вторичной меристемы — феллогена (пробкового камбия). Феллоген, в свою очередь, делясь, откладывает наружу феллему, а внутрь — феллодерму. Образуется перидерма, пробковый слой которой изолирует первичную кору от центрального цилиндра. В результате вся первичная кора отмирает и постепенно сбрасывается; наружным слоем корня становится перидерма. Клетки феллодермы и остатки перицикла в дальнейшем разрастаются и составляют паренхимную зону, которую называют вторичной корой корня (рис. 70).
При развитии запасающей паренхимы главного корня происходит формирование запасающих корней или корнеплодов. Различают корнеплоды:
1. Монокамбиальные (редька, морковь) — закладывается только один слой камбия, а запасные вещества могут накапливаться либо в паренхиме ксилемы (ксилемный тип — редька), либо в паренхиме флоэмы (флоэмный тип — морковь);
2. Поликамбиальные — через определенные промежутки времени происходит заложение нового слоя камбия (свекла).
зоны деления. Это самая верхняя часть корня, представленная клетками апикальной меристемы, которые постоянно делятся. Эти клетки мелкие, изодиаметрические, с тонкими оболочками, густой цитоплазмой и крупными ядрами. Вакуолей в них много, но они мелкие и практически незаметные.
Зону деления снаружи покрывает корневой чехлик. Он имеет вид колпачка и состоит из паренхимных живых клеток, содержащих цитоплазму, ядро, амилопласты с крахмальными зернами и тонкие ослизняющиеся оболочки.
В зоне растяжения клетки обычно прекращают делиться и увеличиваются в размерах. Корень в этой зоне прозрачен, что определяется, прежде всего, образованием крупных вакуолей. Наряду с ростом клеток наблюдается их дифференциация.
Зона всасывания четко заметна благодаря наличию корневых волосков. Здесь большинство клеток уже полностью дифференцированы. По периферии расположены клетки ризодермы. У некоторых растений не все клетки ризодермы способны образовывать волоски. В этом случае выделяют два типа клеток: трихобласты, образующие волоски, и атрихобласты — клетки, выполняющие защитную функцию.
Место, где происходит отмирание корневых волосков, является началом зоны проведения. Она тянется вплоть до корневой шейки и составляет большую часть протяженности корня. На этом участке корня происходит ветвление.
На срезе уже при малом увеличении ясно различаются небольшая внутренняя часть — центральный цилиндр, и наружная первичная кора, покрытая одним слоем клеток с корневыми волосками — ризодермой (эпиблемой).
Наружный слой первичной коры — экзодерма, состоит из плотно сомкнутых многоугольных клеток, стенки которых впоследствии опробковевают и выполняют защитную функцию. Затем расположена основная паренхима (мезодерма), составляющая главную массу первичной коры.
Внутренний слой первичной коры — эндодерма состоит из одного ряда клеток, с утолщенными радиальными и внутренними стенками. Среди этих клеток имеются тонкостенные живые клетки (расположенные почти напротив мелких сосудов ксилемы), называемые пропускными.
Наружный слой центрального цилиндра — перицикл, состоит из одного ряда паренхимных клеток.
Внутренняя часть центрального цилиндра занята полиархным радиальным пучком.
центральный цилиндр с четырьмя лучами первичной ксилемы (тетрархный пучок). Между ними расположены основания четырех крупных открытых коллатеральных проводящих пучков. Эндодерма заметна плохо, так как у ее клеток утолщены лишь радиальные стенки (пятна Каспари). При большом увеличении видно, что клетки тонкостенной паренхимы, лежащей между ксилемой и флоэмой, разделены тангентальными перегородками, а в некоторых местах внутрь от этого слоя заметны только что образовавшиеся и еще не одревесневшие сосуды.
Между ксилемой и флоэмой расположена широкая камбиальная зона, имеющая неровные очертания и состоящая из нескольких рядов довольно мелких клеток таблитчатой формы. Вторичное утолщение связано с заложением и деятельностью камбия. Вторичная ксилема значительно превышает по площади флоэму и лежит ближе к центру. Она представлена крупными сосудами, волокнами и мелкими клетками паренхимы. Вторичная флоэма, находящаяся по периферии камбиальной зоны, представлена ситовидными трубками с простыми горизонтальными ситовидными пластинками, клетками-спутницами и паренхимой. Первичная флоэма расположена на самой периферии пучка, ее ситовидные трубки деформированы.
Между проводящими пучками находятся широкие первичные лубодревесные лучи, образованные межпучковым камбием. Крупные паренхимные клетки, образующие лучи, несколько вытянуты в радиальном направлении.
С поверхности корень тыквы покрыт перидермой.
На поперечном срезе корня моркови под перидермой найти большое кольцо флоэмы. Основная ее масса представлена запасающей паренхимой, а ситовидные трубки образуют небольшие группы.
34.Метаморфозы корней. Часто корни выполняют особые функции, и в связи с этим у них меняется строение. Если корни изменены сравнительно мало, то их морфологическая природа легко устанавливается. В крайних же случаях строение изменено так сильно, что для выяснения морфологической природы требуется специальное исследование, и тогда говорят, что корни метаморфизированы. Под метаморфозом понимают резкое, наследственно закрепленное видоизменение органа, вызванное сменой функций. Видоизменения корней очень разнообразны.
Микориза и сожительство с бактериями. Корни многих растений сожительствуют с почвенными грибами. Корневые окончания и сросшиеся с ними гифы грибов образуют микоризу (дословно «грибокорень»). Высшее растение и гриб извлекают из такого сожительства взаимную пользу, т.е. находятся в состоянии симбиоза. Грибной компонент облегчает корням поглощение воды и минеральных веществ из почвы и, по-видимому, передает им некоторые органические вещества. В свою очередь, гриб получает от высшего растения углеводы и другие питательные вещества. Симбиотические отношения не исключают того, что гриб на некоторых этапах развития угнетает высшее растение и паразитирует на нем, а высшее растение в известные моменты « переваривает» грибные гифы, находящиеся в его клетках. Словом, этот симбиоз можно рассматривать как хорошо отрегулированный паразитизм.
Читайте также: Зил 508 сколько цилиндров
Микотрофное питание (т.е. питание с помощью грибов), видимо, возникло сотни миллионов лет назад у примитивных сухопутных растений, у которых еще не было корней. В настоящее время микотрофия распространена очень широко. Большая часть дикорастуших и культурных трав и деревьев образует микоризу. Некоторые растения, например орхидные, вообще не могут жить без симбиоза с грибами.
Бактериальные клубеньки на корнях бобовых, видимо, представляют собой измененные боковые корни, приспособленные к симбиозу с бактериями. Эти бактерии проникают через корневые волоски внутрь молодых корней и вызывают образование на них клубеньков.
Втягивающие корни могут укорачиваться у своего основания. Так как они прочно срастаются с почвой, то укорочение приводит к втягиванию побега (луковицы, корневища) в почву. Следовательно, втягивающие корни помогают побегам находить наилучшую глубину залегания в почве. Втягивающие корни узнаются по утолщенным основаниям с поперечной морщинистостью.
Запасающие корни обычно утолщены и сильно паренхиматизированы. Запасающая паренхима находится в первичной коре, древесине или сердцевине.
Сильно утолщенные придаточные корни георгина, чистяка, любки называют корневыми шишками.
У многих, чаще всего двулетних, стержнекорневых растений возникает образование, носящее название корнеплода. Морфологическая природа его сложна, так как в его образовании принимают участие и главный корень, и стебель. У моркови почти весь корнеплод, за исключением самой верхней части, составлен корнем. У репы корень образует лишь самую нижнюю часть корнеплода, а его основная часть сформирована гипокотилем (подсемядольным коленом). Можно найти много случаев промежуточного характера.
Воздушные корни образуются у многих тропических эпифитов из семейств орхидных, ароидных, бромелиевых. Эпифиты не паразитируют на деревьях, а только используют их как субстрат, для поднятия вверх, к свету. Воздушные корни орхидей свободно висят в воздухе и приспособлены к поглощению влаги, попадающей на них в виде дождя или росы. На поверхности воздушных корней образуется веламен.
Дыхательные корни хорошо развиты у некоторых тропических мангровых деревьев, обитающих по болотистым побережьям океанов, в полосе приливов и отливов. У авиценнии образуется очень сложная корневая система с дыхательными корнями, вырастающими вертикально вверх. На концах этих корней в покрывающей их перидерме имеется система чечевичек, связанных с аэренхимой. Воздух через эти ткани поступает в подводные органы.
Ходульные корни образуются у деревьев, живущих в тех же мангровых зарослях по берегам океанов, что и авиценния. У этих деревьев, например, образуются ризофоры, от стволов под углом отходят придаточные корни, которые, достигнув грунта, обильно разветвляются. Со временем основания стволов перегнивают, и деревья стоят на этих корнях как на ходулях. Благодаря ходульным корням деревья распределяют свою массу на большую площадь опоры («лыжный эффект»), приобретают устойчивость на зыбком илистом грунте даже во время отливов.
Столбовидные корни (корни-подпорки) особенно замечательны у индийских баньянов. Эти корни закладываются как придаточные на горизонтальных ветвях дерева и свешиваются вниз. Достигнув почвы, они разветвляются в ней и сильно утолщаются, превращаяясь в столбовидные, поддерживающие крону дерева. Такие корни-подпорки позволяют дереву разрастаться в стороны, покрывая площадь до 2500 м2.
Корнеплод — утолщение главного корня и стебля. Он состоит из головки, шейки и корня (рис. 44). Головка — надсемядольная часть растения (эпикотиль),представляет собой стебель с сильно укороченными междоузлиями. Из головки развивается розетка листьев с пазушными почками. Шейка — средняя часть корнеплода; она формируется вследствие разрастания подсе- мядольного колена (гипокоти- ля). У плоских и круглых корнеплодов (редис, репа, брюква и свекла) шейка — основная мясистая часть их. Она не образует корневых ответвлений. У всех длинных корнеплодов (морковь, пастернак, петрушка, некоторые сорта редьки) нижняя часть развивается за счет утолщения главного стержневого корня, вокруг которого образуется развитая всасывающая корневая система.
В связи с характером формирования корнеплода растения, образующие его в основном за счет собственно корня (морковь, петрушка корневая, некоторые сорта редьки и свеклы столовой), нельзя выращивать рассадой или пересаживать, так как при повреждении корня формируются уродливые корнеплоды.
Корнеплодные растения имеют различное соотношение массы листьев и корня: наиболее ценны сорта с малой листовой розеткой. Листья в этом случае обладают высокой продуктивностью, а корнеплоды — наименьшим количеством грубых сосудисто-волокнистых пучков. Поэтому на семена отбирают корнеплоды с маленькой головкой.
При прорастании семян появляется зародышевый корешок, из которого затем развивается главный корень в виде тонкого стержня, направленного вглубь. С появлением первого и второго настоящих листьев в результате деятельности камбиального кольца стержень начинает увеличиваться в диаметре. Первичная кора (покров) отмирает и слущивается. Происходит так называемая линька корня. К этому времени для растений необходимы наиболее благоприятные почвенные и световые условия, что возможно лишь при размещении оптимального числа растений на определенной площади, то есть при оптимальной густоте стояния. Избыточное загущение растений во время линьки приводит к вытягиванию корнеплодов (стеканию) с последующей деформацией, огрубением и недоразвитием их.
На слабоокультуренных почвах с небольшим пахотным горизонтом, а также на почвах, удобренных свежим навозом или на за-
дерненных, длинные корнеплоды петрушки, моркови и пастернака принимают уродливую форму и обычно ветвятся. Ветвление корнеплодов наблюдается также при поражении главного корня. Они ветвятся и при разреженном размещении растений.
У всех цветковых растений корень имеет одинаковое первичное строение. Упрощенная схема первичного строения такова: снаружи корень защищен тонкой кожицей, под которой располагается первичная кора из нескольких слоев, облекающая центральный цилиндр, то есть внутреннюю часть (стержень) всей оси растения. Внешний слой центрального цилиндра (перицикл) дает начало боковым корням. В тонкостенной паренхиме центрального цилиндра возникает деятельный слой камбия. Все ткани, находящиеся внутри камбиального кольца, называют первичной ксилемой, а снаружи (в том числе и перицикл) — первичной флоэмой. С момента возникновения камбиального кольца и с началом его деятельности происходит переход к вторичному строению корня — появляется утолщение, которое приводит к отмиранию и слущиванию первичной коры.
Корнеплод — запасающий орган. У большинства видов корнеплодных растений масса его нарастает за счет деятельности одного камбиального кольца (монокамбиальность). Запасные питательные вещества могут откладываться или в древесинной паренхиме (у редьки, редиса, репы и брюквы), или в коровой паренхиме (у моркови, пастернака, петрушки, сельдерея). У свеклы масса корнеплода нарастает за счет деятельности многих концентрических колец камбия (поликамбиальность). Таким образом, в зависимости от особенностей анатомического строения и вида запасающей ткани различают три типа корнеплода.
Редечный (у редиса, репы, редьки и брюквы) — клетки кам-биального кольца откладывают ксилемные элементы в большем ко-личестве, чем флоэмные, и поэтому основную часть корнеплода составляет древесинная паренхима. Кора же нарастает незначительно, и толщина ее у зрелых корнеплодов не превышает 2. 4 мм.
Морковный — второй тип монокамбиального корнеплода, но с преимущественным развитием коровой паренхимы, покрытой кожицей, и относительно слабым — ксилемы, занимающей внутреннюю часть корнеплода. У моркови толстый слой коры, чаще интенсивно-оранжевой или красной, реже фиолетовой окраски, имеет нежный приятный вкус. Внутренняя же часть — древесина (внутренний стержень) неправильных контуров, бледноок- рашенная, отличается сравнительно грубой, деревянистой консистенцией мякоти.
Свекольный тип (у свеклы столовой, кормовой, сахарной) — корнеплод строением существенно отличается от описанных ранее типов. У свеклы корень и гипокотиль претерпевают первичное, вторичное и третичное изменения. Вначале корень молодого проростка имеет первичное строение, но уже через 10. 12 дней с момента появления настоящих листьев в коре и подсемядольном колене наступают вторичные изменения, обусловленные формированием и деятельностью первого камбиального кольца. Однако такое строение сохраняется недолго и не вызывает существенного утолщения корня и гипокотиля. Вслед за деятельностью первого камбиального кольца начинает делиться кольцо клеток перицикла, интенсивно откладывая внутрь однородные паренхимные клетки. В этой кольцевой паренхиме обособляется второе камбиальное кольцо, с началом деятельности которого рассматривают третичное строение корнеплода свеклы. Наружный слой вторичной коры превращается в третье камбиальное кольцо, и таким же путем очень быстро закладываются следующие камбиальные кольца (всего8. 12), в результате чего образуется поликамбиальный корнеплод. На поперечном срезе корнеплода видны концентрические зоны прироста, состоящие из светлых колец древесины и темноокрашен- ных паренхимных клеток коры. Возникшие многочисленные камбиальные кольца функционируют с различной интенсивностью (молодые кольца наиболее
35. Побе́г (лат. córmus) — один из основных вегетативных органов высших растений, состоящий из стебля с расположенными на нём листьями и почками.
Вегетативный невидоизменённый побег — единый орган растения, состоящий из стебля, листьев и почек, формирующийся из общего массива меристемы (конуса нарастания побега) и обладающие единой проводящей системой. Стебли и листья, являющиеся основными структурными элементами побега — часто рассматриваются как его составные органы, то есть органы второго порядка. Кроме того, обязательная принадлежность побега — почки. Главная внешняя черта, отличающая побег от корня — наличие листьев.
Для побега характерна метамерность, то есть чередование одинаковых частей — метамеров. Листья располагаются на стебле в определённом порядке, в соответствии с типом листорасположения, а участок стебля на уровне отхождения листа называется узлом. Если основание листа полностью окружает стебель, узел называется закрытым, в противном случае — открытым. Промежутки стебля между соседними узлами называют междоузлиями. Каждый повторяемый отрезок побега, который включает в себя узел с находящимся под ним междоузлием, а также расположенные на этом отрезке листья и почки называют метамером. Угол между листом и вышележащим междоузлием называют листовой пазухой, или пазухой листа.
В онтогенезе побег развивается из почек зародыша либо из пазушной или придаточной (адвентивной) почки. Таким образом, почка является зачаточным побегом. При прорастании семени из зародышевой почечки формируется первый побег растения — его главный побег, или побег первого порядка.
Из главного побега формируются боковые побеги, или побеги второго порядка, а при повторении ветвления — третьего порядка и т. д.
Придаточные побеги формируются из придаточных почек.
Так формируется система побегов, представленная главным побегом и боковыми побегами второго и последующего порядков. Система побегов увеличивает общую площадь соприкосновения растения с воздушной средой.
В зависимости от выполняемой функции различают побеги вегетативные, вегетативно-генеративные и генеративные. Вегетативные (невидоизменённые) побеги, состоящие из стебля, листьев и почек и вегетативно-генеративные (частично видоизменённые), состоящие дополнительно из цветка или соцветия, выполняют функции воздушного питания и обеспечивают синтез органических и неорганических веществ. В генеративных (полностью видоизменённых) побегах фотосинтез чаще всего не происходит, зато там образуются спорангии, задача которых сводится к обеспечению размножения растения (к таким побегам относится и цветок).
Побег, на котором образуются цветки, называется цветоносным побегом, или цветоносом (иногда термин «цветонос» понимают в более узком смысле — как участок стебля, на котором находятся цветки).
Почка — это зачаточный побег с очень укороченными междоузлиями. На поперечном срезе почки в ее центре есть зачаточная стебель, окруженный зачаточными листочками, прикрепленные в его нижней части. Зачаточная стебель и зачаточные листья вместе составляют зачаточный побег. Его верхушка называется конусом нарастания. За счет верхушечной почки побег нарастает в высоту, за счет пазушных (их еще называют боковыми) происходит боковое ветвление. Почки, которые закладываются в любом другом месте стебля или на корнях, называют дополнительными. Почки, которые имеют, кроме зародышевого стебля, только зародышевые листки, называют вегетативными, а те, у которых есть и зачаточные цветки или даже соцветия — цветочными (генеративных) почками. Спящие почки — те, которые могут несколько лет находились в состоянии покоя, не развиваясь.
Читайте также: Главный рабочий цилиндр сцепления уаз буханка
Осевая часть почки — зачаточный стебель. От нижней части оси отходят почечные чешуи — это сильно видоизмененные листья. Они кожистые, плотно сомкнуты и нередко пропитаны различными веществами. Почечные чешуи располагаются снаружи почки и защищают находящиеся в ней органы от повреждений.
Выше по оси зачаточного побега расположены зачатки зеленых листьев. В их пазухах находятся очень мелкие, едва различимые в лупу, пазушные почки. Самая верхняя часть зачаточного побега представлена конусом нарастания, благодаря которому он быстро растет после набухания почки.
Вегетативные и генеративные почки. Почка, внутри которой имеется зачаточный стебель с листьями и почками, представляет собой зачаточный вегетативный побег и называется вегетативной. Если в почке есть зачатки цветков, ее называют цветочной или генеративной.
Развертывание почки. Обычно весной при наступлении благоприятных условий почки на побегах набухают. При этом они увеличиваются в размерах, но чешуи у них пока еще остаются плотно сомкнутыми. В это время внутри почки происходит рост всех ее частей, особенно листьев и междоузлий. Вскоре почечные чешуи раздвигаются и появляются зеленые свернутые листочки молодого побега.
После развертывания почек побеги растут быстро, а почечные чешуи почти сразу опадают. Участок побега с листовыми рубцами от опавших почечных чешуй называют почечным кольцом. По числу таких колец можно определить возраст ветки или небольшого дерева и кустарника.
Спящие почки. У деревьев и кустарников не из всех перезимовавших почек образуются побеги. Часть почек остается не раскрытыми. Это спящие почки. Они в течение многих лет могут оставаться живыми и медленно нарастать верхушкой. При повреждении растений (рубка, обрезка, обмерзание) такие почки способны формировать побеги. Спящие почки часто отличаются от обычных большей величиной. Кроме того, новые побеги из спящих почек могут образоваться на старых стволах и пнях д Ветвление — процесс образования новых побегов и характер их взаимного расположения на стебле, многолетней ветви и корневище.
Поскольку побег является осевым органом, у него присутствует апикальная меристема, которая обеспечивает неограниченное нарастание в длину. Такой рост сопровождается более или менее регулярным ветвлением побега. У низших растений в результате ветвления возникает разветвлённый таллом (слоевище), у высших — образуются системы побегов и корней. Ветвление позволяет многократно увеличивать фотосинтетическую поверхность и обеспечивать растение органическими веществами.
У различных растений наблюдается несколько типов ветвления: дихотомическое, моноподиальное, симподиальное. При дихотомическом ветвлении конус нарастания расщепляется надвое. При моноподиальном ветвлении верхушечная почка функционирует в течение всей жизни, образуя главный побег (ось первого порядка), на котором в акропетальной последовательности развиваются оси второго порядка, на них — третьего порядка и т. д. При симподиальном ветвлении одна из верхних пазушных почек образует ось второго порядка, которая растёт в том же направлении, что и ось первого порядка, смещая в сторону её отмирающую часть. Впервые такое деление было предложено немецким ботаником Пфитцером в конце XIX века.
Для Псилотовидных характерно дихотомическое ветвление
Дихотомическое ветвление является наиболее примитивным типом ветвления, наблюдается как у низших растений, так и у некоторых высших растений При дихотомическом ветвлении конус нарастания делится надвое, вновь образующиеся апексы также делятся надвое и так далее.
Различают так называемое изотомическое дихотомическое ветвление (образующиеся ветви равны по размеру) и анизотомическое (при котором образующиеся ветви неравны).
Моноподиальное ветвлениеМоноподиальное ветвление является следующим этапом эволюции ветвления побегов. У растений с моноподиальным типом строения побега верхушечная почка сохраняется на протяжении всей жизни побега. Моноподиальный тип ветвления часто встречается среди голосеменных растений[5], также встречается у многих покрытосеменных (например, у многих видов пальм, а также растений из семейства Орхидные — гастрохилусов, фаленопсисов и других). Некоторые из них имеют единственный вегетативный побег (например, Фаленопсис приятный).
Моноподиальные растения — термин, наиболее часто используемый в описании растений тропической и субтропической флоры, а также в научно-популярной литературе по комнатному и оранжерейному цветоводству.
Моноподиальные растения могут существенно различаться внешне. Среди них есть розеточные, с удлиненным побегом, кустовидные.еревьев.
Камбиальная зона хорошо выражена и представлена довольно мелкими клетками. Площадь сечения ксилемы значительно меньше площади флоэмы. В центре расположена диархная первичная ксилема. Немногочисленные сосуды вторичной ксилемы расположены в виде прерывистых радиальных цепочек в запасающей паренхиме.
На поперечном срезе корня редьки найти ксилему, занимающую наибольшую часть. В центре расположены мелкие сосуды диархной первичной ксилемы. Вторичная ксилема представлена, главным образом, запасающей паренхимой. Небольшие группы сосудов образуют радиальные цепочки. По периферии камбиальной зоны расположено узкое кольцо вторичной флоэмы. Корень покрывает перидерма.
На поперечном срезе корня свеклы найти хорошо выраженные концентрические кольца тканей. Возникновение колец связано с наличием добавочных камбиев, образующихся из перицикла и его производных. Затем рассмотреть в центре корня свеклы диархную первичную ксилему. Между двумя лубодревесными лучами расположены два небольших открытых коллатеральных пучка. Основную массу корня занимает запасающая паренхима, образованная в результате деятельности добавочных камбиев. Многочисленные открытые коллатеральные пучки, представленные небольшим числом сосудов и ситовидных трубок с клетками-спутницами, расположены в виде концентрических колец. Корень покрыт перидермой.
У растений с симподиальным типом строения побега верхушечная почка, закончив развитие, отмирает или дает начало генеративному побегу. После цветения этот побег больше не растет, а у его основания начинает развиваться новый[6]. Структура побега у растений с симподиальным типом ветвления сложнее, чем у растений с моноподиальным типом; симподиальное ветвление является эволюционно более продвинутым типом ветвления.[источник не указан 1808 дней] Слово «симподиальный» образовано от др.-греч. συν- («вместе») и πούς («нога»).
Симподиальное ветвление характерно для многих покрытосеменных растений: например, для лип, ив и многих орхидных.
У орхидных, помимо верхушечных, у части симподиальных орхидей образуются и боковые соцветия, развиваясь из почек, расположенных у основания побега (Пафиния гребенчатая). Часть побега, прижатая к субстрату, называется ризомой. Располагается, как правило, горизонтально и не имеет настоящих листьев, только чешуевидные. Редуцированной, почти неразличимой ризома бывает у многих масдеваллий, дендробиумов и онцидиумов; хорошо различимой и утолщённой — у каттлей и лелий, удлинённой — у бульбофиллюмов и целогин, достигая 10 и более сантиметров. Вертикальная часть побега часто бывает утолщённой, образующей так называемый туберидий, или псевдобульбу. Псевдобульбы могут быть различной формы — от почти сферической до цилиндрической, конусовидной, булавовидной и удлинённой, напоминающей стебли тростника. Псевдобульбы являются запасающими органами.
36.Стебель — это осевая часть побега растений, состоящая из узлов и междоузлий. Стебель выполняет главным образом опорную (механическую) и проводящую функции, обеспечивая наиболее благоприятное для фотосинтеза расположение листьев и двустороннее передвижение растворенных веществ (от корней к листьям, от листьев к другим органам). Кроме того, в стеблях могут откладываться запасные питательные вещества, вода (кактусы); часто стебель является ассимиляционным органом, служит для защиты растения от поедания животными (колючки боярышника, дикой груши), для вегетативного размножения.
Стебель, как и корень, растет в высоту (длину) за счет деления и роста клеток конуса нарастания верхушечной почки.
По степени одревеснения различают стебли травянистые и деревянистые. Травянистый стебель существует обычно один вегетационный период и отличается слабым утолщением, деревянистый — многолетний, способный утолщаться неопределенно долго.
Анатомическое строение стебля соответствует его главным функциям. В нем развита система проводящих тканей, которая связывает воедино все органы растения. С помощью механических тканей стебель поддерживает все надземные органы и выносит листья в благоприятные условия освещения. В стебле имеется система меристем, поддерживающих нарастание его в длину и толщину. Нарастание стебля в толщину обеспечивается вторичной образовательной тканью— камбием, расположенным на границе коры и древесины. У древесных семенных растений камбиальное утолщение может продолжаться долгие годы. Гигантская секвойя образует ствол толщиной свыше 10 м. Кроме того, камбий, расположенный в стебле в виде кольца, обеспечивает определенное постоянство в расположении и составе откладываемых им элементов разных тканей.
Стебель липы имеет типичное для древесных растений строение (рис. 8.10). В центре находится небольшой участок живых тонкостенных клеток сердцевины, в которых накапливаются запасные питательные вещества. Сердцевина окружена толстым слоем древесины (вторичной ксилемы), составляющей около 90% всего объема стебля.
Основные функции типичных надземных стеблей: служить увеличению поверхности растения путем ветвления, образования листьев и наиболее выгодного расположения их; быть посредниками в передвижении веществ между двумя важнейшими органами растений — корнями и листьями; образовывать цветки, при посредстве которых происходит половое размножение растений. Таким образом, роль стебля является сравнительно второстепенной. Во многих случаях (в стволах древесных растений, в подземных стеблях, так называемых корневищах) стебель является местом отложения запасных питательных веществ; у некоторых он функционально заменяет листья (кактусы, спаржа, камыш и др.), служит для защиты (стеблевые колючки), лазанья (стеблевые усики) и т. д
37.Анатомическое строение стебля соответствует его главным функциям: проводящей — в стебле хорошо развита система проводящих тканей, которая связывает все органы растения; опорной — с помощью механических тканей стебель поддерживает все надземные органы и выносит листья в благоприятные условия освещения и ростовой — в стебле, имеется система меристем, поддерживающих нарастание тканей в длину и толщину (верхушечные, боковые» вставочные).
Верхушечная меристема дает начало первичной боковой меристеме — прокамбию — и вставочным меристемам. В результате деятельности первичных меристем формируется первичная структура стебля. Она может сохраняться у некоторых растений длительное время. Вторичная меристема — камбий — формирует, вторичное строение стебля.
Первичная структура. В стебле различают центральный цилиндр (стелу) и первичную кору.
Первичная кора снаружи покрыта эпидермой (покровная ткань), под ней находится хлоренхима (ассимиляционная ткань). Она может образовывать чередующиеся полосы, тянущиеся вдоль стебля, с механическими тканями (колленхимой и склеренхимой). Часто колленхима отсутствует. В первичной коре могут быть неспециализированная паренхима и выделительные ткани.
Центральный цилиндр окружен слоем эндодермы. Основная часть центрального цилиндра занята проводящими тканями (флоэмой и ксилемой), образующими вместе с механической тканью (склеренхимой) сосуд исто-волокнистые пучки. Внутрь от проводящих тканей располагаете» сердцевина, состоящая из неспециализированной паренхимы. Часто в сердцевине образуется воздушная полость.
Вторичная структура — камбий формирует внутрь вторичную ксилему, наружу — вторичную флоэму. Первичная кора отмирает и заменяется вторичной — это совокупность всех вторичных тканей, расположенных снаружи от камбия.
Строение стебля часто сильно отличается от описанной типичной картины. Строение стебля зависит от условий обитания и отражает особенности строения той или иной систематической группы растений.
Дата добавления: 2015-01-19 ; просмотров: 200 ; Нарушение авторских прав
📺 Видео
Внутренне строение корня.Скачать

Вагнеровцы после обороны Бахмута #shortsСкачать

Студенты российского вуза разработали вечный двигатель #вечныйдвигатель #изобретенияСкачать

СТРОГО ПО ЦЕНТРУ !!! БЕЗ СТАНКА И ТОКАРЯ, как просверлить отверстие в болтеСкачать

главный тормозной цилиндрСкачать

Шлифовка внутренней поверхности гильзы цилиндра четвёртой ступени газового компрессораСкачать

Как достать предмет из цилиндра двигателя не разбирая его?Скачать

анатомическое строение корня покрытосеменных растенийСкачать

FIAT Panda ’80 • компактная МЕГА-ПРАКТИЧНОСТЬ 1980-х из ИТАЛИИ • вся ИСТОРИЯ от 1980-х до 2000-х!Скачать

Фрезерование выточек на внутренней поверхности цилиндраСкачать

цилиндр полый с отверстием.Скачать

ПРИЧИНА ГИБЕЛИ ЧЕРЕНКОВ МЕТЕЛЬЧАТОЙ ГОРТЕНЗИИ ДЛЯ МЕНЯ ЯСНА!!Скачать

Кадыров уволил и отправил домойСкачать
